Mosca del cuerno, Haematobia irritans.
Autor: Dr. Elías Alvarez MV para El Federal Veterinario.
- El Insecto, descripción, taxonomía, biología:
La mosca del cuerno es “una pequeña mosca picadora”, mitad del tamaño de la mosca brava, de 2-3mm. Es de color grisáceo y su huésped preferido es el bovino, aunque puede parasitar también al equino, caprino, ovino, mular, caninos y al hombre.
Morfológicamente se diferencia de otras moscas picadoras por el tamaño y forma de sus palpos: son macizos y largos, bien visibles a la lupa y casi tan largos como el órgano picador ( proboscide y estilete ) De esta particular forma de los palpos se supone obtiene su denominación común de mosca de los cuernos.
Cuando está alimentándose despliega sus alas en un ángulo de 60 grados colocando su cabeza hacia el suelo, hecho este característico de la especie.
Taxonomía:
Orden: Díptera, Grupo: Ciclorrafos, Familia: Muscidae, Subfamilia: Stomoxydae, Género: Haematobia, Especie: Irritans, ( en Asia yAustralia, Exigua)
Biología:
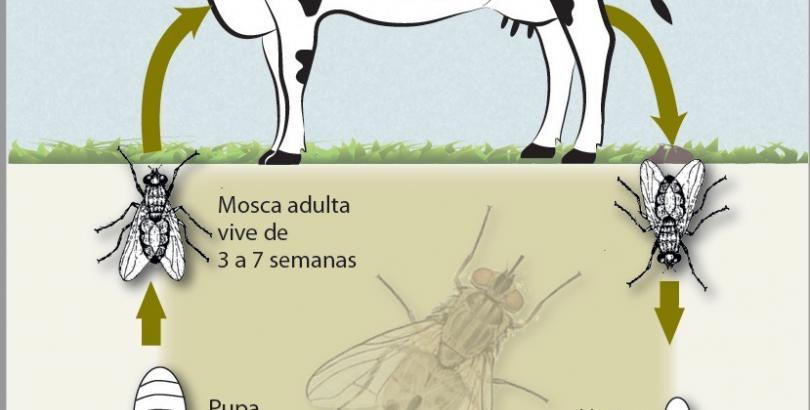
Son ovíparos, con huevos de 1,3 a 1,5 mm. de color marrón a amarillo. El ciclo de vida de huevo a adulto varía según las condiciones de foto período, humedad y temperatura entre 12 a 42 días (mayor calor y temperatura acortan el ciclo).
Permanecen continuamente (6 a 8 semanas) sobre el huésped. Ambos sexos son hematófagos y se aparean sobre el huésped. Cada hembra aova cerca de 20 huevos por vez, y en su vida ovipone hasta 400 huevos.
Son picadoras intermitentes, y se alimentan de 20 a 40 veces al día haciéndolo menos veces el macho.
Bajo determinadas circunstancias abandonan al huésped y vuelan hasta 11 kilómetros para localizar a nuevos hospedadores, este desplazamiento puede responder a formas de resistencia a los insecticidas, resistencia etológica, ver el capítulo “tipos y mecanismos de resistencia”.
El ciclo de la mosca adulta se interrumpe cuando las condiciones de temperatura no le son propicias. Entre 26 y 32 grados los adultos encuentran su condición óptima; si la temperatura baja (15-20 grados) disminuyen su actividad prolongando su ciclo de vida y cuando la temperatura llega a 5 grados, se produce la interrupción del ciclo y comienza la etapa de diapausa, en la bosta, muriendo los adultos.
Oviposición
y fases juveniles:
Cuando la
hembra ha succionado el volumen necesario de sangre, ocurre la ovipostura. El
acto de oviposición es muy rápido: la hembra espera la defecación del bovino,
desciende a la bosta y sobre la boñiga deposita el aove, regresando de inmediato
a la superficie corporal del huésped, el aove solo ocurre sobre bosta fresca,
boñigas mayores a 3 minutos no atraen a las hembras.
De los
huevos eclosionan larvas en 24 horas, estas se internan en la bosta e inician
su alimentación. En 3 a 12 días se producen 2 estadíos más. La supervivencia de
las larvas depende de su habilidad para alejarse de la superficie externa de la
boñiga, buscando zonas de mayor humedad y segura alimentación. Siguiente a la
larva 3, ocurre la pupación, en el límite entre la boñiga y el suelo.
Las pupas son ovoides, elongadas, de color
marrón y de 3 a 4 mm y bien visibles a la lupa. En 2 a 8 días el adulto emerge.
El ciclo juvenil de la mosca ocurre, entonces, exclusivamente en la materia
fecal y dura entre 9 a 12 días; si las condiciones ambientales no son
propicias, la pupa detiene su desarrollo y se inicia la diapausa, sobreviviendo
durante el invierno. Esto determina la variación estacional, hecho de máxima
importancia para la comprensión y diseño de programas de control. En estricta
observación a este desdoblado ciclo biológico, se insistirá repetidamente en
este informe sobre el uso de estrategias eficaces de control.
·
Resistencia:
No obstante
el avance en métodos no químicos para el control de los insectos, el uso de
insecticidas es todavía una herramienta importante para su eficaz control.
El avance
de la resistencia a los insecticidas es un desafío creciente, fruto del notable
resultado del DDT durante la segunda guerra mundial, que hizo imaginar a los
biólogos una derrota de los artrópodos. No obstante la toxicidad e inicio de
resistencia al DDT, se generó una sucesión de nuevos principios que generaron
resistencia cruzada por actuar sobre ”blancos permanentes“ ( neurotoxicidad ),
y ello debido a los escasos puntos de ataque biológico que ofrecen los
artrópodos. A mediados de siglo pasado, la resistencia, más que una amenaza
estratégica fué un desafío para nuevos principios activos. Desconocer la
plasticidad de los artrópodos y los mecanismos defensivos de sus ciclos de vida
y de sus mecanismos genéticos y etológicos nos lleva hoy a replantear la lucha
contra Haematobia irritans, ingresada al país en 1991, pero con una larguísima
migración y consecuente transporte de su memoria genética a los plaguicidas,
ocurrida entre los años 59 y 91.
Tipos y mecanismos
de resistencia:
Existen 2 tipos de resistencia, la fisiológica
o genética, y la etológica o de rechazo.
1)
fisiológica:
1.1: por menor penetración cuticular.
1.2: por
mayor detoxificación metabólica.
1.3: por menor sensibilidad de los receptores
(biofase).
2) etológica:
Se refiere al cambio de actitud o actividad del
insecto enfrentado al insecticida, cambio en el lugar de parasitación, en el
fotoperíodo, etc. Este tipo de resistencia descripta en plagas vegetales y en
Anopheles y Haematobia, Sparks (1989), está siendo revisado. Quisenberry
(1984), encuentra que poblaciones expuestas a caravanas se reubican en ventral,
adjudicando ello a una hiperirritabilidad de la mosca inducida por piretroides.
Byfor (1987), coincide en los estudios y define esta resistencia etológica como
respuesta al estímulo de los insecticidas.
Genética
de la resistencia:
Puede deberse a un solo gen (monogenética) o
bien a varios genes. En la mayoría de los casos, niveles altos de resistencia
son consecuencia de cambios en uno o dos genes, y se producen por duplicación,
por cambios en pares de bases de ADN (alelos) y por cambios en los sitios de
acción y/o en la cantidad de enzimas detoxificadoras.
En estos casos, habiendo un alelo resistente y
otro sensible se expresan tres genotipos:
RESISTENTE RR, denominada resistencia
dominante.
HETEROCIGOTA RS, denominada resistencia
intermedia o codominante.
SENSIBLE SS, denominada resistencia recesiva.
Para la mosca de los cuernos, hay evidencias de
resistencia debida a un gen autosomal, incompletamente recesivo y ligado al
sexo (mosca macho).
Varios autores postulan que alelos de
resistencia para un nuevo insecticida, deben ser muy raros previo a su empleo,
(por una baja tasa de mutaciones en el plantel genético), en consecuencia
solamente la migración de poblaciones portadores de resistencia introduce en
los territorios invadidos por Haematobia, genes RR y/o RS que luego la
imperfección de lmanejo y tratamientos, trastoca a un aumento de la resistencia
y a la pérdida de eficacia sobre poblaciones SS.
·
El aumento de la resistencia:
Para analizar este aspecto debe partirse de la
absoluta dependencia del insecto sobre el huésped, este aspecto como otros no
bien considerados ha creado la crítica situación que proponemos modificar.
La
continua parasitación de la mosca adulta sobre el bovino, la imprescindible
oviposición sobre la reciente defecación y el urgente retorno a la superficie
corporal del bovino, creando cerca de 15 ciclos o generaciones durante la
temporada de parasitación (15 ciclos generaciones x 12 días =180 días de vida
libre) ha creado una condición tal que errores de tratamiento y/o estrategias,
influyen notablemente en la transmisión de información genética que no
deseamos. La mayor o menor cantidad de ciclos anuales tiene influencia en la
multiplicación de la resistencia. Crosby (1991), postula que el retardo en el
surgimiento de la resistencia en el sudoeste de EEUU, se debió a la menor
cantidad de ciclos anuales en esa región. El aumento de ciclos anuales y
errores tácticos en el manejo de plaguicidas mejoran las condiciones para
selección de RR.
Un detallado estudio de la variación de los
ciclos en nuestro país es todavía una aspiración, que nos demostraría
explicaciones a la variante resistencia que estamos encontrando.
·
La influencia de la intensidad en el
uso de insecticidas:
Retardar
o acelerar la formación de resistencia por el empleo de insecticidas depende de
la dosis, la frecuencia de los tratamientos, el poder residual del producto, y
el porcentaje de la población tratada.
Si pudiéramos con altas dosis eliminar el 95%
los heterocigotas RS, si la frecuencia de estos genes fuera baja y hubiera
inmigración al lugar del tratamiento de moscas homocigotas sensibles SS, se
favorecería la mezcla de genes, y disminuiría la posibilidad de formación de
resistencia. Para ello deberíamos esperar que muy pocos sobrevivientes RS, se
mezclaran y reprodujeran con altas cantidades de inmigrantes SS, que el
insecticida actuara rápidamente y por poco tiempo. Esta estrategia lógica, es
sin embargo muy limitada.
El uso de dosis bajas, se ha convertido en un
temible error, por el uso de formulaciones de mala calidad, por formulaciones
caseras, y en EEUU por la combinación del largo efecto de las caravanas y su
“cola de degradación”.
Reducir el número de tratamientos es una de las
más efectivas tácticas para controlar la resistencia. Los intervalos sin tratamiento,
serán intervalos sin contacto de la mosca con el insecticida y permitirán que
fracciones de la población de moscas sin contacto con el insecticida hagan
posible un corrimiento de la biomasa RR a RS e idealmente a SS. Debe intentarse
reducir los tratamientos durante el período de mosca adulta, es decir durante
los aproximadamente 15 ciclos estivales, para que la biomasa aove que entrará
en diapausa traslade a la próxima estación la menor cantidad posible de RR y
eventualmente de RS Esto es la aplicación del concepto “ control integrado de
plagas”.
Sparks (1985) postula un uso intermitente con
insecticidas/productos de escaso poder residual.
Kunz, demostró que 12 ciclos libres de
tratamiento, bajaron el índice de resistencia a la permetrina de 17 a 2,5
veces, pero allí se mantuvo por 20 generaciones, quedando un umbral remanente
de resistencia, que se reactivó al reimplantarse el mismo tratamiento,
(permetrina).
La persistencia de los insecticidas por tiempo
prolongado es una táctica desventajosa por su influencia en el aumento del
proceso selectivo sobre las poblaciones existentes RS y RR, aumentando asi la
formación de resistencia, como se verá en el capítulo “historia de la
resistencia). El uso de caravanas de larga acción conllevó el lamentable
aumento de resistencia en el hemisferio norte, transfiriendo a sucesivas
generaciones y finalmente a la diapausa una mayor biomasa RR.
Sheppard (1978), adjudica el incremento en la
resistencia al efecto insecticida persistente a través de caravanas.
Varios autores han expresado que, la masiva
manifestación de resistencia en EEUU se ha debido a productos con liberación
sostenida, en caravanas. La persistencia de insecticidas, selecciona
permanentemente y por varios ciclos de vida libre genes RR- El uso de caravanas
que seleccionen genes resistentes durante varios ciclos (15 días por ciclo),
constituye entonces una estrategia ,desde el punto de vista de prevención del
incremento de resistencia, inadecuada, más bien un retorno a las prácticas que
ayudaron a generar la alta resistencia en EE.UU, luego llevada por migración .
Sheppard y Joyce (1992) descubrieron los
índices más altos de resistencia, mediante el uso de caravanas con
lambda-cialotrina.
Existe consenso en aceptar que el incremento
y/o aparición de resistencia está estimulado a medida que aumenta el porcentaje
de población expuesta al insecticida.
Tratar fracciones menores de población, crear
zonas sin tratamiento “refugios”, permite que en estos lotes de población no
tratadas, se haga posible el cruzamiento de poblaciones RR y RS con SS.
·
Otras medidas para revertir o
demorar la formación de resistencia:
Uso de mezclas insecticidas: expresa la
exposición simultánea a dos o más insecticidas y/o sinergistas. Se asume que
los homocigotas resistentes RR, sobrevivientes a un tratamiento, difícilmente
son portadores de los alelos de resistencia para otro activo que no haya demostrado
resistencia cruzada. Este efecto se llama de muerte redundante, y diluye la
existencia de genes resistentes, Curtis (1978- 1985), exige además que la
formulación les ofrezca similar persistencia, que la frecuencia de resistencia
sea baja y que una parte de la población se mantenga como refugio, es decir no
tratada, Tabashnik (1990 ).
El uso de fosforados: Los bajos o nulos índices
de resistencia cruzada a piretroides ,Crosby (1991), alentó, el ensayo de
organofosforados; aunque algunos de ellos, ej. stirofos en caravanas, Harvey
(1984), diazinon, McKenzie (1993), ronnel, Burns (1963), mostraron resistencia,
se inició la búsqueda renovada de eficacia de algunos miembros de esta familia
ej. ethion, metil pirimifós, diazinon. El uso de algunos fosforados, debe
tomarse como uno de los pocos y últimos recursos no renovables quimiotácticos,
y evitar cometer con ellos los errores que lamentamos hoy con los piretroides
monodroga.
El uso de ivermectinas tácticas: La ivermectina
es eficaz contra los estadios inmaduros (ciclo larval en la bosta), a la dosis
como endectocida. Su utilización combinada estratégicamente con la dinámica
estacional, y con la frecuencia y efectividad de los plaguicidas es una
herramienta que debe ser cuidadosamente planificada pero debe considerarse el
exceso de uso de esta molécula, hecho que ha acelerado la formación de
resistencia en endoparásitos.
·
Historia de la resistencia:
Según la Organización Mundial de la Salud
(1970), la primera y documentada evidencia de resistencia está registrada en
1959, en Texas y al grupo HCH-dieldrina, luego en 1962, en Louisiana se
describe resistencia a un canfeno clorado (toxafeno). Un año después a un
fosforado (fenclorfos=Ronnel).
Es la propia OMS quien recomienda allí,
establecer sistemas de medición de la resistencia y una aceleración en los
estudios genéticos. Durante los años 82 al 90 el uso de caravanas aumenta los
factores de resistencia, hasta el máximo demostrado de 92.000 veces, en l992 y
a la lambda cialotrina. Esta manifestación explosiva fué atribuida por la
mayoría de investigadores al uso constante y sostenido de las caravanas, y a la
ignorancia de la estrecha dependencia de Haematobia de sus hospedadores. La
migración de poblaciones RR y RS, se vió favorecida a través de las zonas donde
el ecosistema ofrecía variaciones de temperatura y de fotoperíodo ideales para
permitir la mayor cantidad de ciclos y su mayor poder multiplicador. Por ello
en Argentina, quienes estabamos siguiendo la evolución del insecto en su
migración continental norte sur, esperábamos su ingreso por la frontera
noreste. Y así ocurrió en 1991 ( Misiones, Luzuriaga).
Durante los años 91 al 94, coincidieron el
error estratégico de desconsiderar el manejo racional del problema, el error
publicitario de algunas empresas en recomendar la “erradicación” y el desmedido
uso de piretroides de distintas calidades y peor, de formulación casera. Para
1994 (Anziani), no se evidenciaban datos formales de resistencia a
cipermetrina, suponiendo que las poblaciones se expandían y multiplicaban como
RS y RR.
Entre l995 y hoy, se insiste en presionar a las
poblaciones de Haematobia y se inicia el uso de mezclas de piretroides con
sinergizantes (piperonil butóxido=PBO), el uso de PBO, supuso que la vía
metabólica de formación de resistencia (detoxificación) era significativa, pero
Miller (1983), demuestra que el PBO solo reduce un 10% el nivel inicial de
resistencia a permetrina. McDonald (1987), finalmente concluye que el efecto
metabólico detoxificante (sinergismo con BPO), es de escasa importancia en la
resistencia a piretroides.
Se inicia luego el uso de organofosforados,
donde se utilizan el clorpirifós, triclorfon, y ethion. Llegando el Ethion
simultáneamente como caravana, y como Pour on, en tanto diazinón sólo en
caravanas. Los carbamatos, con un modo de acción que asemeja a los
organofosforados, no han sido estudiados como eficaces frente a Haematobia, ni
como alternativas a poblaciones piretroide resistentes. El único carbamato
utlizado es el carbaril, su uso en otras indicaciones fué desalentado, Harrison
(1967).
Última revisión septiembre 2016